
研究内容
当研究室の研究内容と成果
1.ダイオキシンが生殖と次世代の健康に及ぼす影響とその消去法
環境汚染物質であるダイオキシンは、食品汚染を介してヒトの健康を障害する可能性が危惧されています。我々はその真実性、機構、並びに回避方法について、実験動物を用いた基礎研究を行っています。特に、次世代の健康や母の子育て能力を障害する可能性について研究を行っています。
これまでの研究により、ダイオキシンを妊娠ラットに与えると、胎児や初期新生児の黄体形成ホルモン(脳下垂体ホルモンの一種)の生成が障害され(図1、2)、これを起点として成長後にまで継続する性未成熟の固着が起こることを明らかにしました。黄体形成ホルモンの低下はダイオキシン曝露妊娠母ラットにα-リポ酸を与えることにより、完全に回復することも明らかにしています(図3)。α-リポ酸はサプリメントとしても市販されている物質(栄養素)であり、生命維持のために動物体内になくてはならないものの一つです。我々の研究では、ダイオキシンは出産後の母の子育て能力の減退をもたらし得ることも明らかにしつつあります。これは、乳汁分泌や子育て行動の誘起に必要なプロラクチン(これも脳下垂体ホルモンの一種です)がダイオキシンによって減少することが原因ではないかと推定しています。これらの研究から、ダイオキシン等の環境汚染物質が次世代の健康や子育てを障害する機構を明らかにし、またその具体的な解除方法の構築を目指しています。
-代表的論文-
- Yuan, M., et al., Biochem. Pharmacol., 210: 115490-115490 (2023).
- Hattori, Y., et al., Biochem. Pharmacol., 154: 213-221 (2018).
- Koga, T., et al., PLoS ONE, 7: e40322 (2012).
- Takeda, T., et al., J. Biol. Chem., 287: 18440-18450 (2012).
- Mutoh, J., et al., Endocrinology, 147: 927-936 (2006).
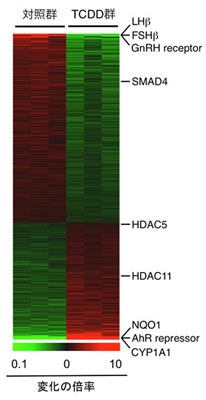 |
図1.ダイオキシンによる胎児・脳下垂体遺伝子の変動:DNAマイクロアレイ解析 最強毒性のダイオキシンである2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)を妊娠ラットに与え、胎児の遺伝子発現を解析。黄体形成ホルモン(luteinizing hormone: LH)のβサブユニットの発現が低下することが読み取れます。緑は明るさが増すほど顕著な低下を示し、逆に、赤は色調の強さに応じて発現が増加する遺伝子であることを示します。変動の程度は図の下に示すカラーグラディエーションを参照。他の解析から、LHβの低下は、histone deacetylase (HDAC) 5 や11の増加によってもたらされることも明らかにしています。対照群(TCDDを投与していない群)とTCDD投与群はそれぞれ3胎児(母は異なる)を解析。 図中には変動が観察されたその他の主な遺伝子も示しています:FSHβ, follicle-stimulating hormone β-subunit; GnRH receptor, gonadotropin-releasing hormone receptor; SMAD4, homolog of Drosophila mothers against decapentaplegic 4; NQO1, NAD(P)H-quinone oxidoreductase; AhR repressor, aryl hydrocarbon receptor repressor; CYP1A1, cytochrome P450 1A1。 |
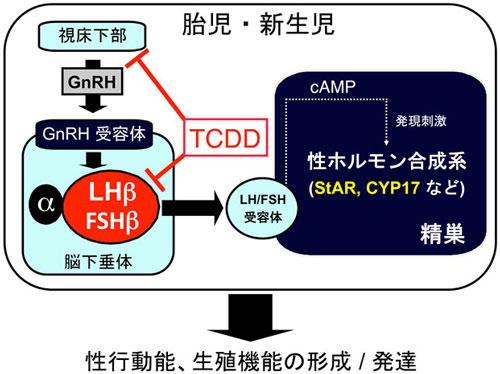 |
図2.TCDDによる脳下垂体・黄体形成ホルモン低下とそれを起点とする性ステロイド合成や性成熟の抑制 |
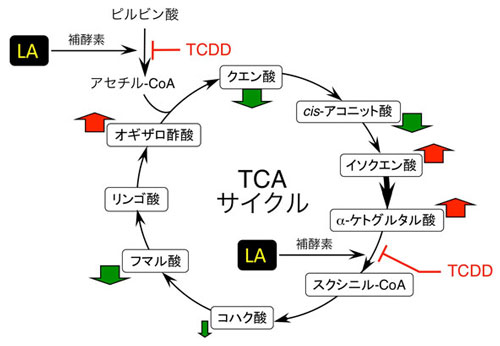 |
図3.TCDDの胎児・視床下部トリカルボン酸(TCA)サイクル抑制とα-リポ酸による回復 TCAサイクルは動物体内でのエネルギー生産に必須な機構です。α-リポ酸(LA)は図中に示す2カ所の反応に必要な物質ですが、これらのステップがTCDDによって抑制されます。これにより、該当ステップは先へ進みにくくなり、前駆体のoxaloacetic acidやα-ketoglutaric acidが蓄積します。体外からLAを補給してやると、TCDD作用が打ち消されて障害が回復すると考えられます。 視床下部は黄体形成ホルモン生産を行う脳下垂体を制御するので、視床下部の活動性を復活させることによって結果として黄体形成ホルモン抑制も回復すると推定しています。 |
2.薬物代謝能力の変動に関与する要因の解明
チトクロムP450(P450)やUDP-グルクロン酸転移酵素(UGT)は、体外異物(通常は身体の中に存在しない物質)の体内での化学変化(薬物代謝と言う)に関わる重要な酵素です。体外異物には医薬品のみならず、食品添加物や食品汚染物質なども含まれます。体外異物は薬物代謝を受けると、多くの場合は不活性化(解毒)されますが、発がん物質等ではかえって有害性を増す場合も知られています。従って、薬物代謝能力の高低は、医薬品の効果や発がん物質による発がんの起こり易さの個人差を規定する重要な要因です。
薬物代謝能の個人差は、P450やUGT等の遺伝子上の変異によって説明できる場合もありますが、他の機構を考えざるを得ない場合も多々あります。我々は、P450とUGTが会合して互いの機能を変化させる独自の学説を提唱し、これが現実に生起することを示すと共にその機構も解明しつつあります(図4、5)。また、UGTについては、脂肪酸誘導体やヌクレオチド類などの生体成分が働きを制御する重要な因子であることを見い出しています(図5)。これらの研究を通して、遺伝子変異では説明できない生体防御機構の変動要因を明らかにしたいと考えています。
-代表的論文-
- Miyauchi, Y., et al., J. Biol. Chem., 301: 110502-110502 (2025).
- Miyauchi, Y., et al., Br. J. Pharmacol., 177: 1077-1089 (2020).
- Ishii, Y., et al., Drug Metab. Dispos., 42, 229-238 (2014).
- Ishii, Y., et al., Drug Metab. Dispos., 40, 2081-2089 (2012).
- Takeda, S., et al., Mol. Pharmacol., 75, 956-964 (2009).
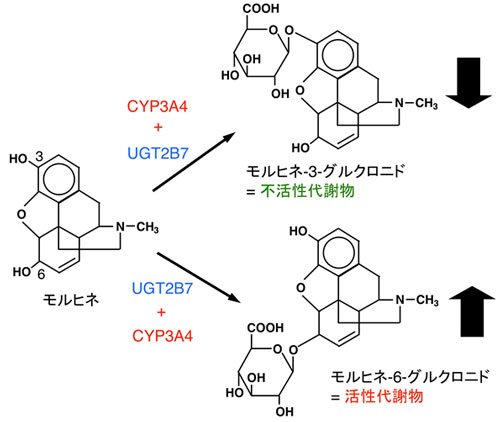 |
図4.UGT2B7によるモルヒネ代謝とCYP3A4による変動 モルヒネはヒト体内でグルクロン酸抱合を受けて、主に3-グルクロニドに変換され鎮痛活性を失います。一部は6?グルクロニドへ変わりますが、こちらはモルヒネよりも強力な鎮痛活性を持っています。これらの2つの代謝は、いずれもUGTの一つのアイソフォームであるUGT2B7が触媒します。我々の研究によって、UGT2B7のモルヒネ代謝は、全く異なる酵素であるCYP3A4が共存すると抑制(3?グルクロニド)および活性化(6?グルクロニド)されることが明らかになりました。 |
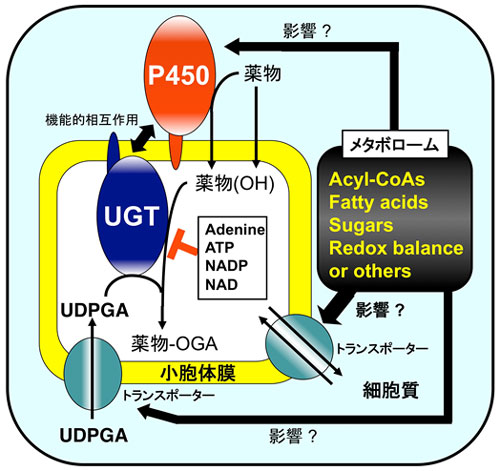 |
図5.CYPやUGTの機能を変動させる要因 我々の研究では、UGTの機能がP450によって変化するのみならず(図4参照)、逆にP450の働きがUGTによって変化することも明らかにしています。また、UGTの機能は種々の生体内成分によって変動することも突き止めています(図に示す成分には予想作用のものも含む)。UDP-グルクロン酸(UDPGA)はUGT反応に必要なグルクロン酸供与体であり、細胞質から反応の場である小胞体内腔へトランスポーターを介して輸送されるものと推定されています。薬物-OGAはグルクロン酸抱合を受けた薬物を指します。 |
3.その他の研究
上記以外にも、薬物代謝酵素の誘導機構(発現が増加する機構)や不正流通薬物の代謝や分析に関する研究も行っています。
-代表的論文-
- Zhao, S., et al., Forensic Toxicol., 43: 256-265 (2025).
- Li, R.S., et al., Sci. Rep., 9: 9621 (2019).
4.生体高分子代謝プロジェクト
オートファジーや細胞内タンパク質輸送など、リソソームの生理機能に関する研究を行っています。
-代表的論文-
- Hirota, Y., et al., Biochem. Biophys. Res. Commun., 566: 9-15 (2021).
- Hirota, Y., et al., Cell. Mol. Life Sci., 66: 2913-2932 (2009).